
Science | 对多种耐药真菌有效的新天然产物的研究

从1928年第一种抗生素青霉素被英国科学家弗莱明发现,到1939年开始被应用于抗细菌感染的治疗,抗生素的发现和应用为人类医学带来了划时代的发展。然而微生物耐药性的问题也随之而来,具有多重耐药性的超级细菌/真菌的出现,对人类健康带来了新的挑战。
目前全球每年约有两百万人死于真菌感染,随着对一线抗真菌药物产生耐药性的病原体的出现,不良状况进一步恶化。耐药菌株如烟曲霉菌(Aspergillus fumigatus)及耳念珠菌(Candida auris)的跨地区传播性及高致死率(50%)给社会健康造成了严重危害。为了应对多重耐药(MDR)病原体的全球威胁,迫切需要新的抗真菌药物。新的抗真菌药物的发展在一定程度上受到了真菌和动物共同进化产生的耐药的影响,并且由于疗效和毒副作用的影响,也限制了靶药物的选择。与其他抗菌药物一样,大多数抗真菌药物来源于天然产物。然而,传统的筛选方法很难筛选到新的药物靶分子,但是基于天然产物的筛选目前是一个新的天然产物来源的动力。总体来说面对微生物耐药性的流行趋势,研究和开发新型药物及治疗手段都迫在眉睫。
近日,美国威斯康星大学麦迪逊分校科学家在Science发表了题为“A marine microbiome antifungal targets urgent-threat drug-resistant fungi”的科学论文,从红树海鞘中发现了对多种耐药真菌有效的新天然产物turbinmicin。
主要的研究方法:利用液相色谱-质谱法(LC-MS)基于代谢组学、基因组学和细菌提取物代谢组抗菌活性阵列分析对收集到的菌株进行筛选
具体的研究思路:
思维导图:


1. 新型抗真菌药物Turbinmicin被发现
研究通过LC-MS代谢组学采集了1482种海鞘放线菌的化学信息,采用HCAPCA(hierarchical cluster analysis principal component analysis)挑选出174种具有化学多样性的菌株,其中WMMC-415(turbinmicin的生产者)在PCA分析中显示出化学差异性(图1C),PCA loadings plot显示了一些导致变异的化合物,包括turbinmicin(图1D)。随后将174种菌株分离、培养,并提取其代谢物,每份菌株提取物采用两步正交色谱法至于96孔板中,采用Candida albicans评估其抗菌效能,筛选出的提取物通过效价强度,MS, NMR确定靶分子的优先顺序,通过过滤已知化合物、关注新型化学骨架,最终鉴定得到新天然产物Turbinmicin。
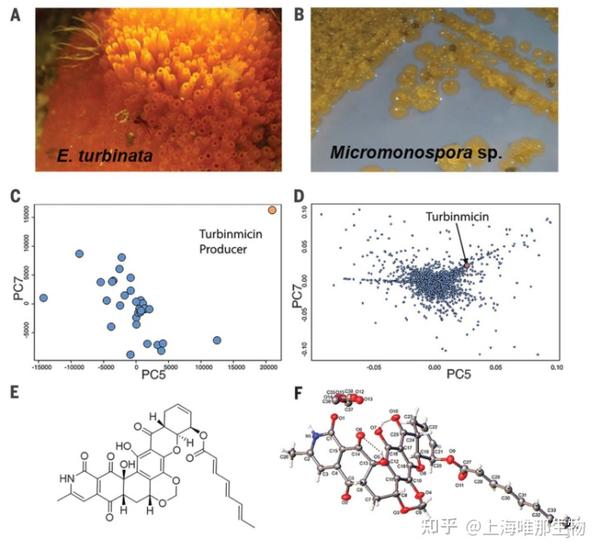

Fig. 1. Turbinmicin discovery from marine microbiome using MDR drug discovery platform.
(A) Turbinmicin-producing strain was isolated from the ascidian Ecteinascidia turbinata. (B) Turbinmicin producing bacterium Micromonospora sp. WMMC-415. (CandD) WMMC-415 was prioritized on the basis of its chemical diversity after HCAPCA processing of a 174-strain library. PCA scores plot (C) revealed WMMC-415’s chemical diversity, and the PCA loadings plot (D) showed a number of diverse metabolites produced by WMMC-415, including turbinmicin; for the purposes of clarity, data in (C) and (D) are from a 30-member subpool of the 174-strain library. (E) Structure of turbinmicin. (F) X-ray ORTEP drawing of turbinmicin shown with 50% probability ellipsoids depicting its absolute configuration
2. Turbinmicin药效及安全性探究
分别采取体外及体内实验对turbinmicin的抗真菌效能进行探究。在体外试验中,turbinmicin对涵盖了多种耐药机制的常见病原真菌(如C. albicans, C. auris, Candida glabrata, Candida tropicalis, A. fumigatus等) 都显示出有效的抗菌性(图2A),能有效抑制多重耐药菌C. auris的生长(图2B)。Turbinmicin浓度超过MIC的1000倍,没有表现出红细胞毒性,这表明该化合物具有广泛的使用范围(图2C),小鼠喂养实验(1mg/Kg~256mg/Kg)中,turbinmicin都未显现出毒副作用,他们继续评估了turbinmicin在嗜中性粒细胞小鼠感染耐药菌株C. auris (strain B11211)中的疗效。结果观察到turbinmicin在感染小鼠中存在剂量依赖性疗效,并且在给药浓度范围内未发现中毒迹象。此外,Turbinmicin 在这些小鼠模型中显示的抗真菌活性与其疗效高度相关,初步验证了其生物安全性.为了进一步评估临床效果,使用美国食品和药物管理局(FDA)的侵袭性念珠菌病标准真菌模型探讨了体内疗效,评估了turbinmicin对高致死率的抗三唑真菌A. fumigatus的疗效。对于体内模型,他们选择中性粒细胞减少和皮质类固醇免疫抑制肺损伤小鼠模型,并通过定量聚合酶链式反应(PCR)进行分析。结果显示turbinmicin对A. fumigatus的剂量依赖性抑制作用与C. auris相似,并且所有的小鼠在turbinmicin治疗期内都表现健康(图2D,2E)。
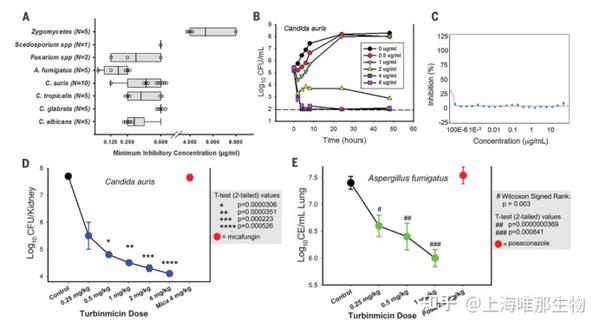

Fig. 2. Turbinmicin displays potent in vitro and in vivo efficacy against multiple MDR fungal pathogens and mammalian safety.
(A) In vitro activity of turbinmicin against 39 fungal isolates. (B) Time-kill curves for turbinmicin against C. auris B11211. Kill curves are generated from data collected at 0, 2, 4, 6,8, 24, and 48 hours after subjection to turbinmicin at concentrations spanning 1 to 16 times the MIC. CFU, colony-forming units. (C) Toxicity of turbinmicin onerythrocytes was performed by hemolysis assay. No hemolytic activity was detected for turbinmicin at all test concentrations. (D) Multidose in vivo experiments with turbinmicin against C. auris B11211 by using a neutropenic, mouse, disseminated candidiasis model (3 mice per dose). The multidose experiments invoked the administration of turbinmicin at doses of 0.25, 0.5, 1, 2, or 4 mg/kg at 6-hour intervals (over a 24-hour period) by intraperitoneal (IP). Mica, micafungin. (E) In vivo multidose experiments with turbinmicin against A. fumigatus F11628 by using a pneumonia model (3 mice per dose). Multidose experiments invoked the administration at doses of 0.25, 0.5, or 1 mg/kg at 6-hour intervals (over a 4-day period) by IP. CE, conidial equivalents; Posa, posaconazole. Statistics handling for both (D) and (E) used pairedttest for normally distributed data and Wilcoxon signed-rank for non-normal data; Pvalues for each data point are indicated in each panel (shaded box).
3. Turbinmicin作用机理的研究
为了破解turbinmicin逆天的抗耐药真菌机制,David R. Andes和Tim S. Bugni团队从酿酒酵母(Saccharomyces cerevisiae)DNA-条形码敲除和敲降库着手。他们使用mRNA干扰(DAmP)敲降文库来降低必需基因的丰度,同时在非必需基因突变的诊断DNA-条形码敲降文库中评估turbinmicin 的效果。化学遗传学分析显示SEC14基因最引人注目。SEC14编码蛋白Sec14p 是一种trans-高尔基体网络的磷脂酰肌醇-磷脂酰胆碱转移蛋白。Sec14p是经过验证的真菌靶点,但迄今为止还从未有批准的抗真菌药物涉及此靶标。非必需敲除文库研究同样支持了Sec14p是turbinmicin 靶标的观点。接着,他们用Transpose NET 构建了一个遗传互作网络,这是一种使用诊断基因集稀疏数据输入网络节点的算法。结果显示SEC14 是该网络的一个“隐藏”中心节点。DAmP技术、诊断非必需遗传数据以及来自单倍剂量不足文库的C. albicans 突变体时间杀灭研究都支持Sec14p作为turbinmicin的主要靶点。另外,他们还直接通过酿酒酵母绿色荧光载运蛋白(GFP)Snc1分泌和内吞模型研究turbinmicin对膜运输的干扰作用。研究发现,在酿酒酵母指数增长期,GFP Snc1 在初生芽质膜中大量积累,并且更温和地定殖于高尔基体和内体膜中。然而,在turbinmicin 的存在下,GFP Snc1不再集中于芽内,而是只存在于高尔基体和内体。
总结:研究先通过采取Chemical Genomics方法推定出turbinmicin可能的作用靶点——Sec14p蛋白(图3A-C),随后采取时间杀菌(C. albicans)实验(图3D),并观察酵母GFP蛋白在不同培养条件下(有无turbinmicin)的转运途径(图3E),证明turbinmicin能通过抑制Sep14蛋白干扰真菌的膜转运功能来发挥抗菌作用。
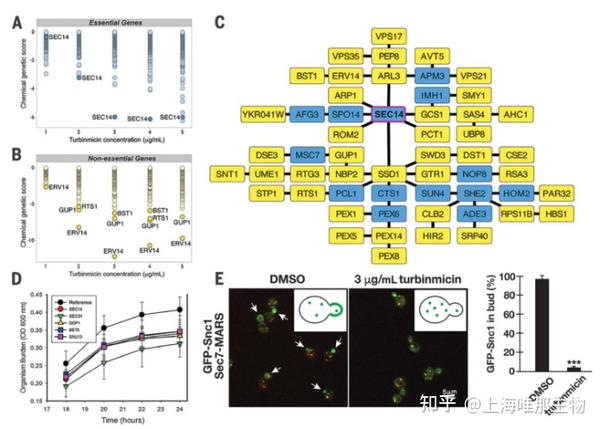

Fig. 3. MOA hypothesis of turbinmicin. Dose-wise chemical-genetic profiles of turbinmicin against the S. cerevisiae DAmP essential gene pool (A) and the diagnostic nonessential gene pool (B). Mean negative chemical-genetic interactions are represented in green [n= 6 except for 2mg/mL turbinmicin in (C),in whichn= 5]. (C) Resulting Transpose NET genetic interaction network built by using sensitive, nonessential gene mutants. Input genes are represented by yellow nodes, and “hidden” nodes are represented in blue. SEC14 was a hidden node and highlighted with a magenta border. (D) Time-kill curve for turbinmicin against wild type and C. albicansmutants. MutantC. albicansSEC14 H1 5C10, DOP1 H1 27E12, BET5 H1 5C5, SUN13 H1 4B5, SEC31 H1(PGA63) 29A6, and SEC31 H1(PGA63) 51H11 were used in this study. OD, optical density. (E) Representative images of GFP-Snc1 localization (green) relative to Golgi membranes (red) in the presence and absence of 3mg/mL turbinmicin (five biological replicates each and at least 100 cells examined per condition; ***P< 0.001 based on attest; error bars are SEM). Spectroscopy with pure turbinmicin (3mg/mL) revealed fluorescence properties distinct and noninterfering with respect to those of the GFP and MARS (mRFPmars). Arrows highlight the distribution of GFP-Snc1 in buds in the absence of turbinmicin (also shown in the graphic). Scale bar, 5mm. DMSO, dimethyl sulfoxide.
文章相关FAQ:
1、什么是LC-MS代谢组学,非靶向代谢组学?
液质联用(LC-MS)又叫液相色谱 - 质谱联用技术,它以液相色谱作为分离系统,质谱为检测系统。样品在质谱部分和流动相分离,被离子化后,经质谱的质量分析器将离子碎片按质量数分开,经检测器得到质谱图。液质联用体现了色谱和质谱优势的互补,将色谱对复杂样品的高分离能力,与 MS 具有高选择性、高灵敏度及能够提供相对分子质量与结构信息的优点结合起来,在药物分析、食品分析和环境分析等许多领域得到了广泛的应用。
非靶向代谢组学(Untargeted metabolomics)是指采用 LC-MS、GC-MS、NMR技术,无偏向性的检测细胞、组织、器官或者生物体内受到刺激或扰动前后所有小分子代谢物(主要是相对分子量1000 Da以内的内源性小分子化合物)的动态变化,并通过生信分析筛选差异代谢物,对差异代谢物进行通路分析,揭示其变化的生理机制。
2、什么是代谢物聚类分析?什么是分层凝聚聚类(Hierarchical Agglomerative Clustering,HAC)
表达模式相似的代谢物通常具有功能相关性,该模块对选择的代谢集中的代谢物进行表达模式聚类分析。依据代谢物在不同样本的表达量信息,计算代谢物或样本的距离,再利用迭代的方法将代谢物或样本进行分类。
适用于:
①实现代谢集中代谢物在各样本中表达的可视化;
② 根据代谢物在各样本中的表达,将代谢物进行聚类,基于表达模式相似的代谢物通常具有功能相关性,可以推断未知代谢物的功能。
分层凝聚聚类原理顾名思义就是要一层一层地进行聚类,可以从下而上地把小的cluster合并聚集,也可以从上而下地将大的cluster进行分割。似乎一般用得比较多的是从下而上地聚集,因此这里我就只介绍这一种。所谓从下而上地合并cluster,具体而言,就是每次找到距离最短的两个cluster,然后进行合并成一个大的cluster,直到全部合并为一个cluster。整个过程就是建立一个树结构,类似于下图。
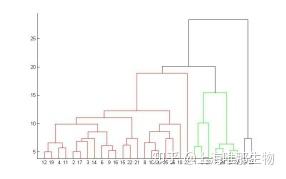
层次聚类最大的优点,就是它一次性地得到了整个聚类的过程,只要得到了上面那样的聚类树,想要分多少个cluster都可以直接根据树结构来得到结果,改变cluster数目不需要再次计算数据点的归属。层次聚类的缺点是计算量比较大,因为要每次都要计算多个cluster内所有数据点的两两距离。另外,由于层次聚类使用的是贪心算法,得到的显然只是局域最优,不一定就是全局最优,这可以通过加入随机效应解决,这就是另外的问题了。
3 什么是主成分分析(Principal components analysis,PCA)
4、什么是NMR核磁共振波谱法(Nuclear Magnetic Resonance spectroscopy,NMR )
5、什么是效价强度(potency)?
6、什么是RNA干扰?
原文链接:
猜你喜欢
SnapGene viewer
微生物基因组公开课回放
1.细菌基因组测序方式:重测序、扫描图、完成图、转录组如何选择
脚本大放送